De novo мутации в гистон-модифицирующих генах как одна из общих причин генетических заболеваний. Мутации de novo при неврологических и психических расстройствах - блог доктора Минутко
Шизофрения - одна из самых загадочных и сложных болезней, причем во многих смыслах. Ее тяжело диагностировать - до сих пор нет консенсуса о том, одно это заболевание или много похожих друг на друга. Ее сложно лечить - сейчас есть лишь препараты, которые подавляют т. н. позитивные симптомы (вроде бреда), но они не помогают возвратить человека к полноценной жизни. Шизофрению сложно исследовать - ни одно другое животное кроме человека ей не болеет, поэтому и моделей для ее изучения почти нет. Шизофрению очень тяжело понять с генетической и эволюционной точки зрения - она полна противоречий, которые биологи пока не могут разрешить. Однако хорошие новости заключаются в том, что в последние годы, наконец, дело вроде бы сдвинулись с мертвой точки. Об истории открытия шизофрении и первых результатах ее изучения нейрофизиологическими методами мы уже . На этот раз речь пойдет о том, как ученые ищут генетические причины возникновения болезни.
Важность этой работы даже не в том, что шизофренией болеет почти каждый сотый человек на планете и прогресс в этой области должен хотя бы радикально упростить диагностику, - даже если создать хорошее лекарство сразу и не получится. Важность генетических исследований в том, что они уже сейчас меняют наши представления о фундаментальных механизмах наследования сложных признаков. Если ученым все-таки удастся понять, как может «прятаться» в нашей ДНК такая сложная болезнь как шизофрения, это будет означать радикальный прорыв в понимании организации генома. И значение такой работы выйдет далеко за пределы клинической психиатрии.
Сначала
немного сырых фактов. Шизофрения
- это тяжелое, хроническое, ведущее к
инвалидности психическое заболевание,
поражающее обычно людей в молодом
возрасте. От нее
страдает около 50 миллионов человек по
всему миру (немногим менее 1% популяции).
Заболевание сопровождается апатией,
безволием, часто галлюцинациями, бредом,
дезорганизацией мышления и речи,
моторными нарушениями. Симптомы обычно
становятся причиной социальной изоляции
и снижения работоспособности. Повышенный
риск суицида у больных шизофренией, а
также сопутствующие соматические
заболевания приводят к тому, что общая
продолжительность жизни у них снижается
на 10-15 лет. Кроме того, больные шизофренией
имеют меньше детей:
мужчины имеют в среднем на 75 процентов, женщины
- на 50 процентов .
Последние полвека стали временем бурного прогресса во многих областях медицины, однако этот прогресс почти не затронул профилактику и лечение шизофрении. Не в последнюю очередь это связано с тем, что мы до сих пор не имеем внятного представления о том, нарушение каких именно биологических процессов является причиной развития заболевания. Такой дефицит понимания привел к тому, что со времени появления на рынке первого антипсихотического препарата хлорпромазина (торговое название: «Аминазин») более 60 лет назад так и не произошло качественного изменения в лечении болезни. Все ныне существующие одобренные для лечения шизофрении антипсихотики (как типичные, включая хлорпромазин, так и атипичные) имеют один и тот же основной механизм действия: они снижают активность дофаминовых рецепторов, что устраняет галлюцинации и бред, но, к сожалению, слабо влияет на негативную симптоматику вроде апатии, безволия, расстройств мышления и т. д. Про побочные эффекты мы даже не упоминаем. Общее разочарование в исследованиях шизофрении проявляется в том, что фармацевтические компании уже давно уменьшают финансирование разработки антипсихотиков , - и это при том, что общее число клинических испытаний только растет. Однако надежда на прояснение причин возникновения шизофрении пришла с довольно неожиданной стороны - она связана с беспрецедентным прогрессом в молекулярной генетике.
Коллективная ответственность
Еще первые исследователи шизофрении заметили, что риск заболеть тесно связан с наличием больных родственников. Попытки установить механизм наследования шизофрении были предприняты почти сразу после переоткрытия законов Менделя, в самом начале XX века. Однако, в отличие от многих других болезней, шизофрения никак не хотела укладывалась в рамки простых менделевских моделей. Несмотря на высокую наследуемость, связать ее с одним или несколькими генами не получалось, поэтому к середине века все большей популярностью стали пользоваться т. н. психогенные теории развития болезни. В согласии с крайне популярным к середине века психоанализом, эти теории объясняли видимую наследуемость шизофрении не генетикой, а особенностями воспитания и нездоровой атмосферой внутри семьи. Появилось даже такое понятие как «шизофреногенные родители».
Однако теория эта, не смотря на свою популярность, прожила недолго. Окончательную точку в вопросе о том, является ли шизофрения наследственной болезнью, поставили психогенетические исследования, проведенные уже в 60-70-е годы. Это были прежде всего близнецовые исследования, а также исследования приемных детей. Суть близнецовых исследований заключается в сравнении вероятностей проявления какого-то признака - в данном случае развития заболевания - у одно- и разнояйцевых близнецов. Поскольку разница в действии среды на близнецов не зависит от того однояйцевые они или разнояйцевые, то различия в этих вероятностях должны происходит главным образом от того, что однояйцевые близнецы генетически идентичны, а разнояйцевые имеют в среднем лишь половину общих вариантов генов.
В случае шизофрении оказалось, что конкордантность однояйцевых близнецов более чем в 3 раза превышает конкордантность разнояйцевых: для первых она составляет приблизительно 50 процентов, а для вторых - менее 15 процентов. Эти слова надо понимать так: если у вас есть страдающий шизофренией однояйцевый брат-близнец, то вы сами заболеете с вероятностью в 50 процентов. Если же вы с братом разнояйцевые близнецы, то и риск заболеть составляет не более 15 процентов. Теоретические расчеты, которые дополнительно учитывают распространенность шизофрении в популяции, дают оценку вклада наследуемости в развитие болезни на уровне 70-80 процентов. Для сравнения, примерно так же наследуется рост и индекс массы тела - признаки, которые всегда считались тесно связанными с генетикой. Кстати, как оказалось позже, столь же высокая наследуемость характерна для трех из четырех остальных основных психических заболеваний: синдрома дефицита внимания и гиперактивности, биполярного расстройства и аутизма.
Результаты близнецовых исследований полностью подтвердились при изучении детей, которые родились у больных шизофренией и были усыновлены в раннем младенчестве здоровыми приемными родителями. Оказалось, что риск заболеть шизофренией у них не снижен по сравнению с детьми, воспитанными своими родителями-шизофрениками, что однозначно указывает на ключевую роль генов в этиологии.
И здесь мы подходим к одной из самых загадочных особенностей шизофрении. Дело в том, что если она так сильно наследуется и при этом очень негативно влияет на приспособленность носителя (напомним, что больные шизофренией оставляют по крайней мере вдвое меньше потомков, чем здоровые люди), то как ей удается сохраняться в популяции по крайней мере на протяжении ? Это противоречие, вокруг которого во многом и происходит главная борьба между разными теориями, получило название «эволюционного парадокса шизофрении»
До недавнего времени ученым было совершенно неясно, какие именно особенности генома больных шизофренией предопределяют развитие болезни. На протяжении десятилетий горячие споры велись даже не о том, какие именно гены изменены у больных шизофренией, а о том, какова общая генетическая «архитектура» болезни.
Имеется ввиду следующее. Геномы отдельных людей очень похожи друг на друга, отличия в среднем составляют менее 0,1 процента нуклеотидов. Некоторые из этих отличительных особенностей генома довольно широко распространены в популяции. Условно считается, что если они встречаются у более чем одного процента людей, их можно называть распространенными вариантами или полиморфизмами. Считается, что такие распространенные варианты появились в геноме человека более 100,000 лет назад, еще до первой эмиграции из Африки предков современных людей, поэтому они присутствуют обычно в большинстве человеческих субпопуляций. Естественно, что для того, чтобы существовать в значительной части популяции на протяжении тысяч поколений большая часть полиморфизмов должна быть не слишком вредна для своих носителей.
Однако в геноме каждого из людей есть и другие генетические особенности,- более молодые и более редкие. Большая часть из них не предоставляет носителям какого-либо преимущества, поэтому их частота в популяции, даже если они фиксируются, остается незначительной. Многие из этих особенностей (или мутаций) имеют более или менее выраженное отрицательное влияение на приспособленность, поэтому они постепенно удаляются негативным отбором. Им взамен в результате непрерывного мутационного процесса появляются другие новые вредные варианты. В сумме частота любой из новых мутаций почти никогда не превышает 0,1 процентов, и такие варианты называют редкими.
Так вот, под архитектурой болезни имеется ввиду то, какие именно генетические варианты - распространенные или редкие, имеющие сильный фенотипический эффект или лишь слегка увеличивающие риск развития болезни, - предопределяют ее появление. Именно вокруг это вопроса до недавнего времени и велись основные споры о генетике шизофрении.
Единственный факт, бесспорно установленный молекулярно-генетическими методами относительно генетики шизофрении за последнюю треть XX века - ее невероятная сложность. Сегодня очевидно, что предрасположенность к болезни определяется изменениями в десятках генов. При этом все предложенные за это время «генетические архитектуры» шизофрении можно объединить в две группы: модель «распространенная болезнь - распространенная изменчивость» («common disease - common variants», CV) и модель «распространенная болезнь - редкие варианты» («common disease - rare variants», RV). Каждая из моделей давала свои объяснения «эволюционного парадокса шизофрении».
RV vs. CV
Согласно модели CV генетическим субстратом шизофрении является некий набор генетических особенностей, полиген, - сродни тому, что определяет наследование количественных признаков вроде роста или массы тела. Такой полиген - это набор полиморфизмов, каждый из которых лишь немного влияет на физиологию (они называются «каузальными», т. к. хоть и не по одиночке, но приводят к развитию болезни). Чтобы поддерживать характерный для шизофрении довольно высокий уровень заболеваемости необходимо, чтобы этот полиген состоял из распространенных вариантов - ведь собрать в одном геноме много редких вариантов очень сложно. Соответственно и каждый человек имеет десятки таких рискованных вариантов в своем геноме. Суммарно все каузальные варианты определяют генетическую предрасположенность (liability) каждого отдельного человека к заболеванию. Предполагается, что для качественных сложных признаков, таких как шизофрения, имеется некое пороговое значение предрасположенности, и заболевание развивается только у тех людей, чья предрасположенность превышает это пороговое значение.
Пороговая модель предрасположенности к заболеванию. Показано нормальное распределение предрасположенности, отложенной по горизонтальной оси. У людей, чья предрасположенность превышает пороговое значение, развивается заболевание.
Впервые такая полигенная модель шизофрении была предложена в 1967 году одним из основателей современной психиатрической генетики Ирвингом Готтесманом, внесшим также значительный вклад в доказательство наследственной природы болезни. С точки зрения приверженцев модели CV сохранение высокой частоты каузальных вариантов шизофрении в популяции на протяжении многих поколений может иметь несколько объяснений. Во-первых, каждый отдельный такой вариант имеет довольно незначительное влияние на фенотип, такие «квази-нейтральные» варианты могут быть невидимы для отбора и оставаться распространенными в популяциях. Особенно это касается популяций с низкой эффективной численностью, где влияние случайности не менее важно, чем давление отбора - к таковым относится и популяция нашего вида.
С другой стороны, выдвигались предположения о присутствии в случае шизофрении т. н. балансирующего отбора, т. е. позитивного влияния «шизофренических полиморфизмов» на здоровых носителей. Это не так уж и сложно представить. Известно, например, что для шизоидных личностей с высокой генетической предрасположенностью к шизофрении (которых много среди близких родственников больных), характерен повышенный уровень творческих способностей, что может слегка увеличивать их адаптацию (это показано уже в нескольких работах). Популяционная генетика допускает такую ситуацию, когда положительный эффект каузальных вариантов у здоровых носителей может перевешивать негативные последствия для тех людей, у которых этих «хороших мутаций» оказалось слишком много, что привело к развитию болезни.
Вторая базовая модель генетической архитектуры шизофрении - модель RV. Она предполагает, что шизофрения - это собирательное понятие и каждый отдельный случай или семья историей заболевания - это отдельная квази-менделевская болезнь, связанная в каждом отдельном случае с уникальными изменениями в геноме. В рамках этой модели каузальные генетические варианты находятся под очень сильным давлением отбора и довольно быстро удаляются из популяции. Но так как в каждом поколении происходит небольшое количество новых мутаций, то между отбором и возникновением каузальных вариантов устанавливается некое равновесие.
С одной стороны, модель RV может объяснить, почему шизофрения очень хорошо наследуется, но ее универсальных генов до сих пор не найдено: ведь в каждой семье наследуются свои собственные каузальные мутации, а универсальных просто нет. С другой стороны, если руководствоваться этой моделью, то приходится признать, что мутации в сотнях разных генов могут приводить к одному и тому же фенотипу. Ведь шизофрения - заболевание распространенное, а возникновение новых мутаций происходит редко. Например, данные по секвенированию троек отец-мать-ребенок показывают, что в каждом поколении на 6 миллиардов нуклеотидов диплоидного генома возникает лишь 70 новых однонуклеотидных замен, из которых в среднем только несколько теоретически могут оказывать какое-либо влияние на фенотип, а мутации других типов - еще более редкое явление.
Тем не менее, некоторые эмпирические данные косвенно подтверждают такую модель генетической архитектуры шизофрении. Например, в начале 90-х годов было обнаружено, что около одного процента всех больных шизофренией имеют микроделецию в одной из областей 22-ой хромосомы. В подавляющем большинстве случаев эта мутация не наследуется от родителей, а происходит de novo в ходе гаметогенеза. Один из 2000 людей рождается с такой микроделецией, приводящей к разнообразным нарушениям в работе организма, названным «синдромом Ди Джорджи». Для страдающих этим синдромом характерны серьезные нарушения когнитивных функций и иммунитета, часто они сопровождаются гипокальциемией, а также проблемами с сердцем и почками. У четверти больных синдромом Ди Джорджи развивается шизофрения. Заманчиво было бы предположить, что и другие случаи шизофрении объясняются сходными генетическими нарушениями с катастрофическими последствиями.
Другим эмпирическим наблюдением косвенно подтверждающим роль de novo мутаций в этиологии шизофрении является связь риска заболеть с возрастом отца. Так, по некоторым данным среди тех, чьим отцам было больше 50 лет на момент рождения, в 3 раза больше больных шизофренией, чем среди тех, чьим отцам было меньше 30. С другой стороны, довольно давно выдвигались гипотезы о связи возраста отца с возникновением de novo мутаций. Такая связь, например, давно установлена для спорадических случаев другой (моногенной) наследственной болезни - ахондроплазии. Эта корреляция совсем недавно была подтверждена вышеупомянутыми данными по секвенированию троек: количество de novo мутаций связано с возрастом отца, но не с возрастом матери. По расчетам ученых от матери ребенок в среднем получает 15 мутаций независимо от ее возраста, а от отца - 25, если ему 20 лет, 55, если ему 35 лет и более 85, если он старше 50. То есть количество de novo мутаций в геноме ребенка увеличивается на две с каждым годом жизни отца.
Казалось, что вместе эти данные довольно ясно указывают на ключевую роль de novo мутаций в этиологии шизофрении. Однако ситуация на самом деле оказалась гораздо сложнее. Уже после разделения двух основных теорий, на протяжении десятилетий генетика шизофрении находилась в стагнации. Не было получено почти никаких достоверных воспроизводимых данных в пользу одной из них. Ни об общей генетической архитектуре болезни, ни о конкретных вариантах, влияющих на риск развития заболевания. Резкий скачок произошел за последние 7 лет и он связан прежде всего с технологическими прорывами.
В поисках генов
Секвенирование первого генома человека, последующее усовершенствование технологий секвенирования, а затем появление и повсеместное внедрение высокопроизводительного секвенирования позволили наконец получить более или менее полное представление о структуре генетической вариабельности в человеческой популяции. Эта новая информация сразу стала использоваться для полномасштабного поиска генетических детерминант предрасположенности к тем или иным заболеваниям, в том числе и к шизофрении.
Строятся подобные исследования примерно так. Сначала собирается выборка неродственных больных людей (cases) и примерно такая же по размеру выборка неродственных здоровых индивидуумов (controls). У всех этих людей определяется наличие тех или иных генетических вариантов - как раз в последние 10 лет у исследователей появилась возможность определять их на уровне целых геномов. Затем производится сравнение частоты встречаемости каждого из определенных вариантов между группами больных людей и группой контроля. Если при этом удается найти статистически достоверное обогащение того или иного варианта у носителей, его называют ассоциацией. Таким образом среди необъятного числа существующих генетических вариантов находятся те, которые связаны с развитием болезни.
Важной величиной, характеризующей эффект ассоциированного с болезнью варианта, является OD (odds ratio, отношение рисков), которое определяется как отношение шансов заболеть у носителей данного варианта по сравнению с теми людьми, у которых он отсутствует. Если величина OD варианта равна 10, это означает следующее. Если взять случайную группу носителей варианта и равную ей группу людей, у которых данный вариант отсутствует, окажется, что в первой группе больных будет в 10 раз больше, чем во второй. При этом чем ближе OD к единице у данного варианта, тем бóльшая выборка нужна для того, чтобы достоверно подтвердить то, ассоциация действительно существует, - что это генетический вариант действительно влияет на развитие болезни.
Подобные работы позволили к настоящему времени обнаружить по всему геному более десятка субмикроскопических делеций и дупликаций , ассоциированных с шизофренией (их называют CNV - copy number variations, одна из CNV как раз вызывает уже известный нам синдром Ди Джорджи). Для обнаруженных CNV, вызывающих шизофрению, OD колеблется в интервале от 4 до 60. Это высокие значения, однако из-за чрезвычайной редкости даже суммарно все они объясняют только очень небольшую часть наследуемости шизофрении в популяции. Что же отвечает за развитие болезни у всех остальных?
После сравнительно неудачных попыток найти такие CNV, которые бы вызывали развитие болезни не в нескольких редких случаях, а у значительной части популяции, сторонники «мутационной» модели возлагали большие надежды на другой тип экспериментов. В них сравнивают у больных шизофренией и здоровых контролей не наличие массивных генетических перестроек, а полные последовательности геномов или экзомов (совокупностей всех кодирующих белки последовательностей). Такие данные, получаемые с использованием высокопроизводительного секвенирования, позволяют находить редкие и уникальные генетические особенности, которые невозможно обнаружить другими методами.
Удешевление секвенирования сделало в последние годы возможным эксперименты такого типа на довольно больших выборках - включающих в последних работах несколько тысяч больных и столько же здоровых контролей. Каков результат? Увы, пока удалось обнаружить лишь один ген, редкие мутации в котором достоверно ассоциированы с шизофренией - это ген SETD1A , кодирующий один из важных белков, участвующих в регуляции транскрипции. Как и в случае с CNV, проблема тут та же самая: мутации в гене SETD1A не могут объяснять сколько-нибудь значимой части наследуемости шизофрении из-за того, что они просто очень редкие.

Связь распространенности ассоциированных генетических вариантов (по горизонтальной оси) и их влияния на риск развития шизофрении (OR). На основном графике красными треугольниками показаны некоторые из обнаруженных к настоящему времени CNV, ассоциированные с болезнью, синими кружками – SNP по данным GWAS. Во врезе в тех же координатах представлены области редких и частых генетических вариантов.
Есть указания на то, что существуют и другие редкие и уникальные варианты, которые влияют на предрасположенность к шизофрении. И дальнейшее увеличение выборок в экспериментах с использованием секвенирования должно помочь отыскать некоторые из них. Однако, несмотря на то, что исследование редких вариантов еще может принести некоторое количество ценной информации (особенно эта информация будет важна для создания клеточных и животных моделей шизофрении), большинство ученых в настоящее время сходятся во мнении, что редкие варианты играют лишь второстепенную роль в наследуемости шизофрении, а модель CV намного лучше описывает генетическую архитектуру болезни. Убежденность в верности CV модели пришла прежде всего с развитием исследований типа GWAS, о которых мы подробно расскажем во второй части. Коротко говоря, исследования такого типа позволили обнаружить ту самую распространенную генетическую изменчивость, описывающую значительную долю наследуемости шизофрении, существование которой предсказывалось моделью CV.
Дополнительным подтверждением CV модели для шизофрении является связь между уровнем генетической предрасположенности к шизофрении и так называемыми расстройствами шизофренического спектра. Еще ранние исследователи шизофрении заметили, что среди родственников больных шизофренией часто встречаются не только другие больные шизофренией, но и «эксцентрические» личности со странностями характера и симптоматикой сходной с шизофренической, но выраженной менее ярко. Впоследствии подобные наблюдения привели к концепции, согласно которой существует целый набор болезней, для которых характерны более или менее выраженные нарушения в восприятии реальности. Эта группа болезней получила название расстройства шизофренического спектра. Помимо различных форм шизофрении к ним относят бредовые расстройства, шизотипическое, параноидное и шизоидное расстройства личности, шизоаффективное расстройство и некоторые другие патологии. Готтесман, предлагая свою полигенную модель шизофрении, предположил, что у людей с субпороговыми значениями предрасположенности к болезни могут развиваться другие патологии шизофренического спектра, причем тяжесть заболевания коррелирует с уровнем предрасположенности.
Если эта гипотеза верна, логично предположить, что генетические варианты, обнаруженные как ассоциированные с шизофренией, будут обогащены и среди людей, страдающих расстройствами шизофренического спектра. Для оценки генетической предрасположенности каждого отдельного человека используется специальная величина, называемая уровнем полигенного риска (polygenic risk score). Уровень полигенного риска учитывает суммарные вклад всех идентифицированных в GWAS распространенных рискованных вариантов, имеющихся в геноме данного человека, в предрасположенность к болезни. Оказалось, что, как и предсказывала модель CV, значения уровня полигенного риска коррелируют не только с самой шизофренией (что тривиально), но и с другими болезнями шизофренического спектра, причем тяжелым типам расстройств соответствуют более высокие уровни полигенного риска.
И все-таки остается одна проблема - феномен «старых отцов». Если большая часть эмпирических данных подтверждает полигенную модель шизофрении, как согласовать с ней давно известную связь между возрастом отцовства и риском детей заболеть шизофренией?
Некогда было выдвинуто изящное объяснение этого феномена с точки зрения модели CV. Предполагалось, что позднее отцовство и шизофрения не являются соответственно причиной и следствием, а представляют собой два следствия общей причины, а именно генетической предрасположенности поздних отцов к шизофрении. С одной стороны, высокий уровень предрасположенности к шизофрении может коррелировать у здоровых мужчин с более поздним отцовством. С другой стороны, очевидно, что высокая предрасположенность отца предопределяет повышенную вероятность того, что его дети заболеют шизофренией. Выходит, что мы можем иметь дело с двум независимыми корелляциями, а значит накопление мутаций в предшественниках сперматозоидов у мужчин можетпочти никак не влиять на развитие шизофрении у их потомков. Недавно полученные результаты моделирования , учитывающего эпидемиологические данные, а также свежие молекулярные данные по частоте de novo мутаций, хорошо согласуются именно с таким объяснением феномена «старых отцов».
Таким образом, в настоящий момент можно считать, что убедительных аргументов в пользу «мутационной» RV модели шизофрении уже почти не осталось. А значит ключ к этиологии болезни лежит в том, какой именно набор распространенных полиморфизмов вызывает шизофрению в соответсвии с CV-моделью. Тому, как этот набор ищут генетики и что им уже удалось обнаружить, будет посвящена вторая часть нашей истории.
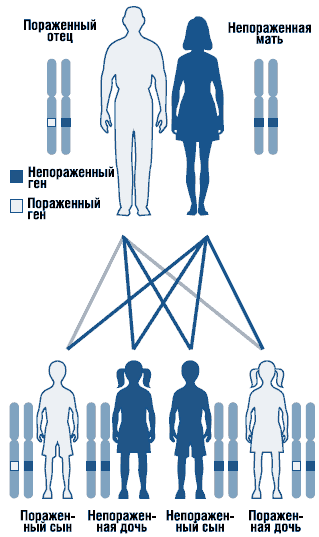
Аркадий ГоловГЛАВА 5 ИЗМЕНЧИВОСТЬ ОРГАНИЗМА
ГЛАВА 5 ИЗМЕНЧИВОСТЬ ОРГАНИЗМА
Общие данные
Изменчивость организма есть изменчивость его генома, обусловливающая генотипические и фенотипические различия человека и вызывающая эволюционное разнообразие его генотипов и фенотипов (см. главы 2 и 3).
Внутриутробное развитие зародыша, эмбриона, плода, дальнейшее постнатальное развитие организма человека (младенчество, детство, отрочество, юность, взрослая жизнь, старение и смерть) осуществляются в соответствии с генетической программой онтогенеза, сформировавшейся при слиянии материнского и отцовского геномов (см. главы 2 и 12).
В ходе онтогенеза геном организма индивида и закодированная в нем информация подвергаются непрерывным преобразованиям под действием факторов окружающей среды. Возникшие в геноме изменения могут передаваться из поколения в поколение, обусловливая изменчивость признаков и фенотипа организма у потомков.
В начале XX в. немецкий зоолог В. Хэкер выделил направление генетики, посвященное изучению связей и взаимоотношений между генотипами и фенотипами и анализу их изменчивости, и назвал его феногенетикой.
В настоящее время феногенетики выделяют два класса изменчивости: ненаследственную (или модификационную), которая не передается из поколения в поколение, и наследственную, которая передается из поколения в поколение.
В свою очередь, наследственная изменчивость также бывает двух классов: комбинативная (рекомбинационная) и мутационная. Изменчивость первого класса определяют три механизма: случайные встречи гамет при оплодотворении; кроссинговер, или мейотическая рекомбинация (обмен равными участками между гомологичными хромосомами в профазе первого деления мейоза); независимое расхождение гомологичных хромосом к полюсам деления при образовании дочерних клеток в ходе митоза и мейоза. Изменчивость второго
класса обусловлена точковыми, хромосомными и геномными мутациями (см. ниже).
Последовательно рассмотрим различные классы и типы изменчивости организма на разных этапах его индивидуального развития.
Изменчивость при оплодотворении гамет и начало функционирования генома зародившегося организма
Материнский и отцовский геномы не могут функционировать отдельно друг от друга.
Только двух родительских генома, объединившиеся в зиготе, обеспечивают зарождение молекулярной жизни, появление нового качественного состояния - одного из свойств биологической материи.
На рис. 23 отражены результаты взаимодействия двух родительских геномов при оплодотворении гамет.
Согласно формуле оплодотворения: зигота = яйцеклетка + сперматозоид, начало развития зиготы - это момент формирования двойного (диплоидного) при встрече двух гаплоидных наборов родительских гамет. Именно тогда зарождается молекулярная жизнь и запускается цепь последовательных реакций на основе сначала экспрессии генов генотипа зиготы, а затем генотипов появившихся из нее дочерних соматических клеток. Отдельные гены и группы генов в составе генотипов всех клеток организма начинают «включаться» и «выключаться» в ходе реализации генетической программы онтогенеза.
Ведущая роль в происходящих событиях принадлежит яйцеклетке, имеющей в ядре и цитоплазме все необходимые для зарожде-
Рис. 23. Результаты взаимодействия двух родительских геномов при оплодотворении гамет (рисунки по www.bio.1september.ru; www.bio.fizteh.ru; www. vetfac.nsau.edu.ru соответственно)
ния и продолжения жизни структурные и функциональные компоненты ядра и цитоплазмы (суть биологического матриархата). Сперматозоид же содержит ДНК и не содержит компонентов цитоплазмы. Проникнув в яйцеклетку, ДНК сперматозоида вступает в контакт с ее ДНК, и тем самым в зиготе «включается» функционирующий в течение всей жизни организма главный молекулярный механизм: ДНК-ДНК взаимодействие двух родительских геномов. Строго говоря, активизируется генотип, представленный примерно равными частями нуклеотидных последовательностей ДНК материнского и отцовского происхождения (без учета мтДНК цитоплазмы). Упростим сказанное: начало молекулярной жизни в зиготе - нарушение постоянства внутренней среды яйцеклетки (ее гомеостаза), а вся последующая молекулярная жизнь многоклеточного организма - стремление восстановить подверженный действию факторов среды гомеостаз или баланс между двумя противоположными состояниями: стабильностью с одной стороны и изменчивостью с другой. Таковы причинно-следственные связи, определяющие возникновение и непрерывность молекулярной жизни организма в ходе онтогенеза.
Теперь обратим внимание на результаты и значение изменчивости генома организма как продукта эволюции. Сначала рассмотрим вопрос об уникальности генотипа зиготы или клетки-родоначальницы всех клеток, тканей, органов и систем организма.
Само оплодотворение происходит случайно: одну женскую гамету оплодотворяет только одна мужская гамета из 200-300 млн сперматозоидов, содержащихся в эякуляте мужчины. Очевидно, что каждую яйцеклетку и каждый сперматозоид отличают друг от друга многие генотипические и фенотипические признаки: наличие измененных или неизмененных по составу и комбинациям генов (результаты комбинативной изменчивости), разные сиквенсы нуклеотидных последовательностей ДНК, разные размеры, форма, функциональная активность (подвижность), зрелость гамет и др. Именно эти отличия позволяют говорить об уникальности генома любой гаметы и, следовательно, генотипа зиготы и всего организма: случайность оплодотворения гамет обеспечивает появление на свет генетически уникального организма индивида.
Иными словами, молекулярная жизнь человека (как и жизнь биологического существа вообще) - «дар судьбы» или, если угодно, «божественный дар», ибо вместо данного индивида с одинаковой
вероятностью могли родиться генетически иные - его родные братья и сестры.
Теперь продолжим наши рассуждения о балансе между стабильностью и изменчивостью наследственного материала. В широком смысле, поддержание такого баланса - это одновременное сохранение и изменение (преобразование) стабильности наследственного материала под действием внутренних (гомеостаз) и внешних средовых факторов (норма реакции). Гомеостаз зависит от генотипа, обусловленного слиянием двух геномов (см. рис. 23). Норма реакции определяется взаимодействием генотипа с факторами окружающей среды.
Норма и диапазон реакции
Специфический способ реакции организма в ответ на действие факторов окружающей среды называется нормой реакции. Именно гены и генотип ответственны за развитие и диапазон модификаций отдельных признаков и фенотипа всего организма. Вместе с тем, в фенотипе реализуются далеко не все возможности генотипа, т.е. фенотип - частный (для индивида) случай реализации генотипа в конкретных условиях окружающей среды. Поэтому, например, между монозиготными близнецами, имеющими полностью идентичные генотипы (100% общих генов), выявляются заметные фенотипические различия, если близнецы растут в разных условиях окружающей среды.
Норма реакции бывает узкой или широкой. В первом случае стабильность отдельного признака (фенотипа) сохраняется практически вне зависимости от влияния окружающей среды. Примерами генов с узкой нормой реакции или непластичных генов служат гены, кодирующие синтез антигенов групп крови, окраску глаз, курчавость волос и др. Их действие одинаково при любых (совместимых с жизнью) внешних условиях. Во втором случае стабильность отдельного признака (фенотипа) изменяется в зависимости от влияния окружающей среды. Пример генов с широкой нормой реакции или пластичных генов - гены, контролирующие количество эритроцитов крови (разное у лиц, поднимающихся в гору, и лиц, спускающихся с горы). Другой пример широкой нормы реакции - изменение окраски кожных покровов (загар), связанный с интенсивностью и временем воздействия на организм ультрафиолетового облучения.
Говоря о диапазоне реакции, следует иметь в виду фенотипические различия, проявляющиеся у индивида (его генотипа) в зависимости от
«обедненных» или «обогащенных» условий окружающей среды, в которых находится организм. Согласно определению И.И. Шмальгаузена (1946), «наследуются не признаки, как таковые, а норма их реакции на изменения условий существования организмов».
Таким образом, норма и диапазон реакции - это пределы генотипической и фенотипической изменчивости организма при изменении условий окружающей среды.
Следует также отметить, что из внутренних факторов, оказывающих влияние на фенотипическое проявление генов и генотипа, определенное значение имеют пол и возраст индивида.
Внешние и внутренние факторы, определяющие развитие признаков и фенотипов, входят в указанные в главе три группы основных факторов, среди которых гены и генотип, механизмы межмолекулярных (ДНК-ДНКовых) и межгенных взаимодействий между родительскими геномами и факторы окружающей среды.
Безусловно, основой приспособления организма к условиям окружающей среды (основой онтогенеза) является его генотип. В частности, индивиды с генотипами, не обеспечивающими подавление отрицательного действия патологических генов и факторов среды, оставляют меньше потомков, чем те индивиды, у которых нежелательные эффекты подавляются.
Вероятно, что в генотипы более жизнеспособных организмов включены специальные гены (гены-модификаторы), подавляющие действие «вредных» генов таким образом, что вместо них доминантными становятся аллели нормального типа.
НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ
Говоря о ненаследственной изменчивости генетического материала, снова рассмотрим пример широкой нормы реакции - изменение окраски кожных покровов под действием ультрафиолетового излучения. «Загар» ведь не передается из поколения в поколение, т.е. не наследуется, хотя в его возникновении участвуют пластичные гены.
Точно так же не наследуются результаты травм, рубцовые изменения тканей и слизистых оболочек при ожоговой болезни, обморожениях, отравлениях и многие другие признаки, вызванные действием исключительно факторов среды. Вместе с тем, следует подчеркнуть: ненаследственные изменения или модификации связаны с наслед-
ственными свойствами данного организма, ибо образуются на фоне конкретного генотипа в конкретных условиях окружающей среды.
Наследственная комбинативная изменчивость
Как сказано в начале главы, кроме механизма случайных встреч гамет при оплодотворении, комбинативная изменчивость включает механизмы кроссинговера в первом делении мейоза и независимого расхождения хромосом к полюсам деления при образовании дочерних клеток в ходе митоза и мейоза (см. главу 9).
Кроссинговер в первом делении мейоза
За счет механизма кроссинговера сцепление генов с хромосомой регулярно нарушается в профазе первого деления мейоза в результате перемешивания между собой (обмена) генов отцовского и материнского происхождения (рис. 24).
В начале XX в. при открытии кроссинговера Т.Х. Морган и его ученики предположили: кроссинговер между двумя генами может происходить не только в одной, но и в двух, трех (соответственно двойной и тройной кроссинговер) и большем количестве точек. Отмечалось подавление кроссинговера в участках, непосредственно примыкающих к точкам обмена; такое подавление назвали интерференцией.
В конечном итоге подсчитали: на один мужской мейоз приходится от 39 до 64 хиазм или рекомбинаций, а на один женский мейоз - до 100 хиазм.
 Рис. 24.
Схема кроссинговера в первом делении мейоза (по Шевченко В.А. и соавт., 2004):
Рис. 24.
Схема кроссинговера в первом делении мейоза (по Шевченко В.А. и соавт., 2004):
a - сестринские хроматиды гомологичных хромосом до начала мейоза; б - они же во время пахитены (видна их спирализация); в - они же во время диплотены и диакинеза (стрелки указывают на места кроссинговера-хиазмы, или участки обмена)
В результате сделали вывод: сцепление генов с хромосомами постоянно нарушается в ходе кроссинговера.
Факторы, влияющие на кроссинговер
Кроссинговер - один из регулярных генетических процессов в организме, контролируемый многими генами как непосредственно, так и через физиологическое состояние клеток в ходе мейоза и даже митоза.
К факторам, влияющим на кроссинговер, относятся:
Гомо- и гетерогаметный пол (речь идет о митотическом кроссинговере у самцов и самок таких эукариот, как дрозофила и тутовый шелкопряд); так, у дрозофилы кроссинговер протекает нормально; у тутового шелкопряда - либо тоже нормально, либо отсутствует; у человека следует обратить внимание на смешанный («третий») пол и конкретно на роль кроссинговера при аномалиях развития пола при мужском и женском гермафродитизме (см. главу 16);
Структура хроматина; на частоту кроссинговера в разных участках хромосом влияет распределение гетерохроматиновых (прицентромерные и теломерные участки) и эухроматиновых районов; в частности, в прицентромерных и теломерных участках частота кроссинговера снижена, и расстояние между генами, определяемое по частоте кроссинговера, может не соответствовать фактическому;
Функциональное состояние организма; по мере увеличения возраста меняется степень спирализации хромосом и скорость клеточного деления;
Генотип; в его составе выделены гены, увеличивающие или уменьшающие частоту кроссинговера; «запиратели» последнего - хромосомные перестройки (инверсии и транслокации), затрудняющие нормальную конъюгацию хромосом в зиготене;
Экзогенные факторы: воздействие температуры, ионизирующей радиации и концентрированных растворов солей, химические мутагены, лекарства и гормоны, как правило, повышающие частоту кроссинговера.
По частоте мейотического и митотического кроссинговера и СХО порой судят о мутагенном действии лекарств, канцерогенов, антибиотиков и других химических соединений.
Неравный кроссинговер
В редких случаях в ходе кроссинговера наблюдаются разрывы в несимметричных точках сестринских хроматид, и они обменива-
ются между собой неравными участками - это неравный кроссинговер.
Вместе с тем, описаны случаи, когда в ходе митоза наблюдается митотическая конъюгация (неправильное спаривание) гомологичных хромосом и рекомбинация происходит между несестринскими хроматидами. Такое явление получило название генной конверсии.
Значение данного механизма трудно переоценить. Например, в результате неправильного спаривания гомологичных хромосом по фланкирующим повторам может произойти удвоение (дупликация) или утрата (делеция) участка хромосомы, содержащего ген РМР22, что обусловит развитие наследственной аутосомно-доминантной моторно-сенсорной нейропатии Шарко-Мари-Тус.
Неравный кроссинговер - один из механизмов возникновения мутаций. Например, периферический белок миелин кодируется геном РМР22, расположенным в хромосоме 17 и имеющим длину около 1,5 млн н.п. Этот ген фланкируется двумя гомологичными повторами длиной около 30 тыс. н.п. (повторы расположены на флангах гена).
Особенно много мутаций в результате неравного кроссинговера происходит в псевдогенах. Тогда либо фрагмент одного аллеля переносится в другой аллель, либо фрагмент псевдогена - в ген. Например, подобная мутация отмечается при переносе последовательности псевдогена в ген 21-гидроксилазы (CYP21B) при адреногенитальном синдроме или врожденной гиперплазии коры надпочечников (см. главы 14 и 22).
Кроме того, за счет рекомбинаций в ходе неравного кроссинговера могут образовываться множественные аллельные формы генов, кодирующих антигены HLA класса I.
Независимое расхождение гомологических хромосом к полюсам деления при образовании дочерних клеток в ходе митоза и мейоза
Благодаря процессу репликации, предшествующему митозу соматической клетки, общее количество нуклеотидных последовательностей ДНК увеличивается вдвое. Формирование одной пары гомологичных хромосом происходит из двух отцовских и двух материнских хромосом. При распределении этих четырех хромосом в две дочерние клетки каждая из клеток получит одну отцовскую и одну материнскую хромосомы (для каждой пары хромосомного набора), однако какую именно из двух, первую или вторую, неизвестно. Имеет место
случайный характер распределения гомологичных хромосом. Легко подсчитать: за счет различных комбинаций 23 пар хромосом общее количество дочерних клеток составит 2 23 , или более 8 млн (8 χ 10 6) вариантов комбинаций хромосом и расположенных на них генов. Следовательно, при случайном характере распределения хромосом в дочерние клетки каждая из них будет иметь свой уникальный кариотип и генотип (свой вариант комбинации хромосом и сцепленных с ними генов соответственно). Следует отметить и возможность патологического варианта распределения хромосом в дочерние клетки. Например, попадание в одну из двух дочерних клеток только одной (отцовской или материнской по происхождению) Х-хромосомы приведет к моносомии (синдром Шерешевского-Тернера, кариотип 45, ХО), попадание трех одинаковых аутосом приведет к трисомии (синдромы Дауна, 47,XY,+21; Патау, 47,ХХ,+13 и Эдвадса, 47,ХХ,+18; см. также главу 2).
Как отмечено в главе 5, в одну дочернюю клетку могут одновременно попасть две отцовские или две материнские по происхождению хромосомы - это однородительская изодисомия по конкретной паре хромосом: синдромы Сильвера-Рассела (две материнские хромосомы 7), Беквитта-Видемана (две отцовские хромосомы 11), Ангельмана (две отцовские хромосомы 15), Прадера-Вилли (две материнские хромосомы 15). В целом объем нарушений распределения хромосом достигает 1% всех хромосомных нарушений у человека. Эти нарушения имеют большое эволюционное значение, ибо создают популяционное разнообразие кариотипов, генотипов и фенотипов человека. Причем каждый патологический вариант является уникальным продуктом эволюции.
В результате второго мейотического деления образуются 4 дочерние клетки. В каждую из них отойдет по одной либо материнской, либо отцовской хромосоме из всех 23 хромосом.
Чтобы избежать возможных ошибок в наших дальнейших расчетах, примем за правило: в результате второго мейотического деления также образуется 8 млн вариантов мужских гамет и 8 млн вариантов женских гамет. Тогда ответ на вопрос, каков общий объем вариантов комбинаций хромосом и расположенных на них генов при встрече двух гамет, следующий: 2 46 или 64 χ 10 12 , т.е. 64 триллиона.
Образование такого (теоретически возможного) количества генотипов при встрече двух гамет наглядно объясняет смысл гетерогенности генотипов.
Значение комбинативной изменчивости
Комбинативная изменчивость важна не только для гетерогенности и уникальности наследственного материала, но и для восстановления (репарации) стабильности молекулы ДНК при повреждении ее обеих нитей. Примером служит образование одноцепочечной бреши ДНК напротив нерепарированного повреждения. Появившаяся брешь не может быть безошибочно исправлена без привлечения к репарации нормальной нити ДНК.
Мутационная изменчивость
Наряду с уникальностью и гетерогенностью генотипов и фенотипов в результате комбинативной изменчивости огромный вклад в вариабельность генома и фенома человека вносит наследственная мутационная изменчивость и обусловленная ею генетическая гетерогенность.
Вариации нуклеотидных последовательностей ДНК чисто условно можно разделить на мутации и генетический полиморфизм (см. главу 2). Вместе с тем, если гетерогенность генотипов - это постоянные (нормальные) характеристики вариабельности генома, то мутационная изменчивость - это, как правило, его патология.
В пользу патологической вариабельности генома свидетельствуют, например, неравный кроссинговер, неправильное расхождение хромосом к полюсам деления при образовании дочерних клеток, наличие генетических компаундов и аллельных серий. Иными словами, наследственная комбинативная и мутационная изменчивость проявляется у человека значительным генотипическим и фенотипическим разнообразием.
Уточним терминологию и рассмотрим общие вопросы теории мутаций.
ОБЩИЕ ВОПРОСЫ ТЕОРИИ МУТАЦИЙ
Мутация есть изменение структурной организации, количества и/или функционирования наследственного материала и синтезируемых им белков. Это понятие впервые предложил Гуго де Фриз
в 1901-1903 гг. в своей работе «Мутационная теория», где описал основные свойства мутаций. Они:
Возникают внезапно;
Передаются из поколения в поколение;
Наследуются по доминантному типу (проявляются у гетерозигот и гомозигот) и рецессивному типу (проявляются у гомозигот);
Не имеют направленности («мутирует» любой локус, вызывая незначительные изменения или затрагивая жизненно важные признаки);
По фенотипическому проявлению бывают вредными (большинство мутаций), полезными (крайне редко) или безразличными;
Возникают в соматических и половых клетках.
Кроме того, одни и те же мутации могут возникнуть повторно.
Мутационный процесс или мутагенез, есть непрерывно идущий процесс формирования мутаций под действием мутагенов - факторов среды, повреждающих наследственный материал.
Впервые теория непрерывно идущего мутагенеза предложена в 1889 г. русским ученым из Петербургского университета С.И. Коржинским в его книге «Гетерогенезис и эволюция».
Как принято считать в настоящее время, мутации способны проявиться спонтанно, без видимых внешних причин, но под влиянием внутренних условий в клетке и организме - это спонтанные мутации или спонтанный мутагенез.
Мутации, вызванные искусственно путем воздействия внешних факторов физической, химической или биологической природы, - это индуцированные мутации, или индуцированный мутагенез.
Наиболее часто встречающиеся мутации называются мажорными мутациями (например, мутации в генах миодистрофии Дюшенна- Беккера, муковисцидоза, серповидноклеточной анемии, фенилкетонурии и др.). Сейчас созданы коммерческие наборы, позволяющие выявлять в автоматическом режиме наиболее важные из них.
Вновь возникшие мутации называются новыми мутациями или мутациями de novo. Например, к ним относятся мутации, лежащие в основе ряда аутосомно-доминантных болезней, таких, как ахондроплазия (10% случаев заболевания - семейные формы), нейрофиброматоз Реклингаузена I типа (50-70% - семейные формы), болезнь Альцгеймера, хорея Гентингтона.
Мутации от нормального состояния гена (признака) к патологическому состоянию называются прямыми.
Мутации от патологического состояния гена (признака) к нормальному состоянию называются обратными или реверсиями.
Впервые способность к реверсии установлена в 1935 г. Н.В. Тимофеевым-Рессовским.
Последующие мутации в гене, подавляющие первичный мутантный фенотип, называются супрессорными. Супрессия может быть внутригенной (восстанавливает функциональную активность белка; аминокислота не соответствует исходной, т.е. истинной обратимости нет) и внегенной (изменяется структура тРНК, в результате чего мутантная тРНК включает в полипептид другую аминокислоту вместо кодируемой дефектным триплетом).
Мутации в соматических клетках называются соматическими мутациями. Они формируют патологические клеточные клоны (совокупность патологических клеток) и в случае одновременного присутствия в организме нормальных и патологических клеток приводят к клеточному мозаицизму (например, при наследственной остеодистрофии Олбрайта экспрессивность заболевания зависит от количества аномальных клеток).
Соматические мутации могут быть как семейными, так и спорадическими (несемейными). Они лежат в основе развития злокачественных новообразований и процессов преждевременного старения.
Ранее считалось аксиомой, что соматические мутации не наследуются. В последние же годы была доказана передача из поколения в поколение наследственной предрасположенности 90% мультифакториальных форм и 10% моногенных форм рака, проявляющихся мутациями в соматических клетках.
Мутации в половых клетках называются герминативными мутациями. Считается, что они встречаются реже соматических мутаций, лежат в основе всех наследственных и некоторых врожденных болезней, передаются из поколения в поколение и также могут быть семейными и спорадическими. Наиболее изученная область общего мутагенеза - физический и, в частности, радиационный мутагенез. Любые источники ионизирующей радиации пагубны для здоровья человека, они, как правило, оказывают мощное мутагенное, тератогенное и канцерогенное воздействие. Мутагенный эффект однократной дозы облучения гораздо выше, чем при хроническом облучении; доза облучения в 10 рад удваивает частоту мутаций у человека. Доказано: ионизирующее излучение способно вызвать мутации, приводящие
к наследственным (врожденным) и онкологическим болезням, а ультрафиолетовое - индуцировать ошибки репликации ДНК.
Большую опасность представляет химический мутагенез. В мире существует около 7 млн химических соединений. В народном хозяйстве, на производстве и в быту постоянно применяются примерно 50-60 тыс. химических веществ. Ежегодно внедряются в практику около одной тысячи новых соединений. Из них 10% в состоянии индуцировать мутации. Таковы гербициды и пестициды (доля мутагенов среди них достигает 50%), а также ряд лекарственных препаратов (некоторые антибиотики, синтетические гормоны, цитостатики и др.).
Существует еще биологический мутагенез. К биологическим мутагенам относятся: чужеродные белки вакцин и сывороток, вирусы (ветряная оспа, коревая краснуха, полиомиелит, простой герпес, СПИД, энцефалит) и ДНК, экзогенные факторы (неполноценное белковое питание), соединения гистамина и его производные, стероидные гормоны (эндогенные факторы). Усиливают действие внешних мутагенов комутагены (токсины).
В истории генетики немало примеров значения связей между генами и признаками. Один из них - классификация мутаций в зависимости от их фенотипического эффекта.
Классификация мутаций в зависимости от их фенотипического эффекта
Такую классификацию мутаций впервые предложил в 1932 г. Г. Мёллер. Согласно классификации были выделены:
Аморфные мутации. Это состояние, при котором признак, контролируемый патологическим аллелем, не проявляется, так как патологический аллель не активен по сравнению с нормальным аллелем. К таким мутациям относятся ген альбинизма (11q14.1) и около 3000 аутосомно-рецессивных заболеваний;
Антиморфные мутации. В этом случае значение признака, контролируемого патологическим аллелем, противоположно значению признака, контролируемого нормальным аллелем. К таким мутациям относятся гены около 5-6 тыс. аутосомно-доминантных заболеваний;
Гиперморфные мутации. В случае такой мутации признак, контролируемый патологическим аллелем, выражен сильнее признака, контролируемого нормальным аллелем. Пример - гете-
розиготные носители генов болезней нестабильности генома (см. главу 10). Их число составляет около 3% населения Земли (почти 195 млн человек), а количество самих заболеваний достигает 100 нозологий. Среди этих заболеваний: анемия Фанкони, атаксиятелеангиэктазия, пигментная ксеродерма, синдром Блума, прогероидные синдромы, многие формы рака и др. При этом частота рака у гетерозиготных носителей генов этих заболеваний в 3-5 раз выше, чем в норме, а у самих больных (гомозигот по этим генам) частота рака в десятки раз выше, чем в норме.
Гипоморфные мутации. Это состояние, при котором проявление признака, контролируемого патологическим аллелем, ослаблено по сравнению с признаком, контролируемым нормальным аллелем. К таким мутациям относятся мутации генов синтеза пигментов (1q31; 6p21.2; 7p15-q13; 8q12.1; 17p13.3; 17q25; 19q13; Xp21.2; Xp21.3; Xp22), а также более 3000 форм аутосомно-рецессивных заболеваний.
Неоморфные мутации. О такой мутации говорят, когда признак, контролируемый патологическим аллелем, будет иного (нового) качества по сравнению с признаком, контролируемым нормальным аллелем. Пример: синтез новых иммуноглобулинов в ответ на проникновение в организм чужеродных антигенов.
Говоря о непреходящем значении классификации Г. Мёллера, следует отметить, что спустя 60 лет после ее публикации фенотипические эффекты точковых мутаций были разделены на разные классы в зависимости от оказываемого ими воздействия на структуру белкового продукта гена и/или уровень его экспрессии.
В частности, нобелевский лауреат Виктор Маккьюсик (1992) выделил мутации, изменяющие последовательность аминокислот в белке. Оказалось, что именно они отвечают за проявление 50-60% случаев моногенных болезней, а остальные мутации (40-50% случаев) приходятся на долю мутаций, затрагивающих экспрессию генов.
Изменение аминокислотного состава белка проявляется в патологическом фенотипе, например, в случаях метгемоглобинемии или серповидноклеточной анемии, обусловленной мутациями бетаглобинового гена. В свою очередь, были выделены мутации, затрагивающие нормальную экспрессию гена. Они приводят к изменению количества генного продукта и проявляются фенотипами, связанными с недостаточностью того или иного белка, например,
в случаях гемолитической анемии, обусловленной мутациями генов, локализованных на аутосомах: 9q34.3 (дефицит аденилаткиназы); 12p13.1 (дефицит триозофосфатизомеразы); 21q22.2 (дефицит фосфофруктокиназы).
Классификация мутаций В. Маккьюсика (1992) - это, безусловно, новое поколение классификаций. Вместе с тем, накануне ее публикации широкое признание получила классификация мутаций в зависимости от уровня организации наследственного материала.
Классификация мутаций в зависимости от уровня организации наследственного материала
Классификация включает следующее.
Точковые мутации (нарушение структуры гена в разных его точках).
Строго говоря, к точковым мутациям относятся изменения нуклеотидов (оснований) одного гена, ведущие к изменению количества и качества синтезируемых ими белковых продуктов. Изменения оснований - это их замены, вставки, перемещения или выпадения, которые можно объяснить мутациями в регуляторных областях генов (промотор, сайт полиаденилирования), а также в кодирующих и некодирующих областях генов (экзоны и интроны, сайты сплайсинга). Замены оснований ведут к появлению трех типов мутантных кодонов: миссенс-мутации, нейтральные мутации и нонсенс-мутации.
Точковые мутации наследуются как простые менделевские признаки. Они часто встречаются: 1 случай на 200-2000 рождений - это первичный гемохроматоз, неполипозный рак толстой кишки, синдром Мартина-Белл и муковисцидоз.
Точковые мутации, встречающиеся крайне редко (1:1 500 000), - это тяжелый комбинированный иммунодефицит (ТКИД) в результате дефицита аденозиндезаминазы. Иногда точковые мутации формируются не при воздействии мутагенов, а как ошибки репликации ДНК. При этом их частота не превышает 1:10 5 -1:10 10 , так как они исправляются с помощью репарационных систем клетки почти на
Структурные мутации или аберрации хромосом (нарушают структуру хромосом и приводят к формированию новых групп сцепления генов). Это делеции (утраты), дупликации (удвоения), транслокации (перемещения), инверсии (поворот на 180°) или инсерции (вставки) наследственного материала. Такие мутации характерны для сомати-
ческих клеток (включая стволовые клетки). Их частота составляет 1 на 1700 клеточных делений.
Известен ряд синдромов, обусловленных структурными мутациями. Наиболее известные примеры: синдром «кошачьего крика» (кариотип: 46,ХХ,5р-), синдром Вольфа-Хиршхорна (46,ХХ, 4р-), транслокационная форма синдрома Дауна (кариотип: 47, ХУ, t (14;21)).
Другой пример - это лейкемии. При них происходит нарушение экспрессии гена в результате так называемого разделения (транслокация между структурной частью гена и его промоторной областью), и, следовательно, нарушается синтез белка.
Геномные (численные) мутации - нарушение числа хромосом или их частей (ведут к появлению новых геномов или их частей путем добавления или утраты целых хромосом или их частей). Происхождение этих мутаций обусловлено нерасхождением хромосом в митозе или мейозе.
В первом случае - это анеуплоиды, тетраплоиды с неразделенной цитоплазмой, полиплоиды, имеющие по 6, 8, 10 пар хромосом и более.
Во втором случае - это неразделение парных хромосом, участвующих в формировании гамет (моносомии, трисомии) или оплодотворение одной яйцеклетки двумя сперматозоидами (диспермия или триплоидный зародыш).
Их типичные примеры уже не раз приводились - это синдром Шерешевского-Тернера (45,ХО), синдром Клайнфельтера (47,ХХУ), регулярная трисомия при синдроме Дауна (47,ХХ, +21).
Неврологические и психические расстройства составляют 13% глобального бремени болезней, что напрямую затрагивает более 450 миллионов человек во всем мире. Распространенность этих расстройств, вероятно, будет продолжать расти в результате увеличения продолжительности жизни населения. К сожалению, почти половина пациентов с шизофренией в настоящее время не получают соответствующей медицинской помощи, отчасти потому, что ранние симптомы шизофрении часто смешиваются с теми, которые наблюдаются при других психических расстройствах (например, психотическая депрессия или биполярное расстройство). Другие расстройства, такие как синдром Ретта (RTT) и нейрофиброматоз типа II (NF2), требуют многодисциплинарного подхода и лечения в специализированных медицинских центрах. Кроме того, большинство этих расстройств являются сложными, в результате взаимодействия генетических и экологических факторов.
На основании данных двойных исследований наследуемость некоторых психических расстройств является высокой. Это относится к аутизму и шизофрении, с наследуемыми факторами порядка 90% и 80% соответственно. Тем не менее, эти заболевания также часто встречаются в виде отдельных случаев, причем может быть только один пострадавший ребенок, рожденный незатронутыми родителями без семейной истории болезни. Одним из возможных объяснений этого феномена является появление мутаций de de novo , где мутации происходят во время сперматогенеза или оогенеза (мутации зародышевой линии) и поэтому присутствуют у пациента, но не обнаруживаются у незатронутого родителя. Этот генетический механизм в последнее время был в центре внимания при объяснении части генетической основы нарушений развити яневрной системы.
Учитывая тот факт, что человеческий геном, по оценкам, содержит примерно 22 333 гена, можно говорить о том, что более 17 800 генов выражены в мозге человека. Мутации, затрагивающие почти любой из этих генов, в сочетании с факторами окружающей среды, могут способствовать появлению неврологических и психических расстройств головного мозга. Недавние исследования выявили ряд причинно-следственных мутаций в генах и показали значительную роль, которую играет генетика в неврологических и психических расстройствах. В этих исследованиях было показано участие редких (<1% частоты) точечных мутаций и вариаций числа копий (CNVs, то есть геномных делеций или дублирования от> 1 кб до нескольких Мб), которые могут возникать в свободных от гена областях, или которые могут повлиять на один ген, или включать в себя непрерывный набор генов в генетической этиологии аутизма, шизофрении, интеллектуальной недостаточности, синдрома дефицита внимания и других нейропсихиатрических расстройств.
Уже давно известно, что неврологические и психические расстройства появляются одних и тех же в семьях, что предполагает наследуемость с основным генетическим компонентом болезни. Для некоторых неврологических расстройств, таких как NF2 или RTT, генетическая причина была идентифицирована. Однако для подавляющего большинства неврологических и психических расстройств, таких как шизофрения, аутизм, биполярное расстройство и синдром беспокойных ног, генетические причины остаются в основном неизвестными. Последние разработки в технологиях секвенирования ДНК открыли новые возможности для нашего понимания генетических механизмов, лежащих в основе этих расстройств. Используя массивные параллельные платформы для секвенирования ДНК (также называемые «следующим поколением») в одном образце (эксперименте) можно искать мутации во всех генах генома человека.
Известно значение De Novo мутаций (то есть, приобретенные мутации в потомстве) при психических расстройствах, таких как умственная отсталость (ID), аутизм и шизофрения. Действительно, при многих недавних исследованиях генома, анализ геномов пораженных лиц и сравнение их с геномами родителей показали, что редкие кодирующие и некодирующие вариации de novo достоверно связаны с риском возникновения аутизма и шизофрении. Было выдвинуто предположение, что большое количество новых случаев этих расстройств частично вызвано мутациями de novo, которые могут компенсировать аллельные потери из-за сильно сниженной репродуктивной способности, тем самым поддерживая высокие частоты этих заболеваний. Удивительно, что мутации de novo довольно распространены (в порядке 100 новых мутаций на ребенка), причем лишь несколько (в порядке по одному на каждого ребенка) в кодирующих областях.
Мутации de novo за пределами областей кодирования, например, в промоторных, интронных или межгенных областях, также могут быть связаны с болезнью. Однако задача состоит в том, чтобы определить, какая из этих мутаций является патогенной.
Несколько основных линий доказательства должны быть приняты во внимание при оценке патогенности наблюдаемой De Novo мутациями: De Novo скорость мутации, функция гена, влияние мутации и клинические корреляции. Основные вопросы сейчас можно сформулировать следующим образом: сколько генов будет вовлечено в неврологические и психические расстройства? Какие конкретные генные пути задействованы? Каковы последствия мутаций de novo для генетического консультирования? На эти вопросы необходимо найти ответы для улучшения диагностики и разработки методов лечения.
Роль мутаций de novo в заболеваниях человека хорошо известна, особенно в области онкологической генетики и доминирующих менделевских расстройств, таких как синдромы Кабуки и Шинцеля-Гиедона (Kabuki , Schinzel-Giedion syndromes). Оба этих синдрома характеризуются выраженной интеллектуальной недостаточностью и врожденными лицевыми аномалиями, причем в последнее время было установлено, что они вызваны мутациями de novo в генах MLL2 и SETBP1 , соответственно. В последнее время исследования Sanders et al ., Neale et al ., O"Roak et al . подтвердили вклад De Novo мутации в этиологии аутизма. Каждое исследование идентифицировало список мутаций de novo, присутствующих у пробандах, но только несколько генов были идентифицированы с несколькими de novo (CHD8, SCN2A, KATNAL2 и NTNG1 ). Протеин-взаимодействие и анализ на основе путей из этих исследований показали значительную взаимосвязь и общий биологический путь между генами, несущими мутации de novo в случаях аутизма. Протеиновые сети, вовлеченные в ремоделирование хроматина, убиквитинирование и развитие нейронов, были идентифицированы как потенциальные мишени для генов восприимчивости к аутизму. Наконец, эти исследования показывают, что 1000 или более генов могут быть интерпретированы как те, в которых могут произойти, как проникающие мутации, способствующие появлению аутизма.
Технологические достижения в секвенировании ДНК по сути сделали революцию в изучении генетических вариаций в геноме человека и позволили идентифицировать многие типы мутаций, включая замены одной пары оснований, вставки / делеции, CNV, инверсии и повторные экспансии, а также рассматриваемые, как соматические и зародышевые мутации. Было показано, что все эти типы мутаций играют определенную роль в заболеваниях человека. Одиночные нуклеотидные мутации, по-видимому, в основном относятся к "отцовскому происхождению", тогда как делеции могут быть в основном "материнского происхождения". Это можно объяснить различиями между мужским и женским гаметогенезом. Например, при исследовании нейрофиброматоза 16 из 21 мутации состояли из делеций материнского происхождения, а 9 из 11 точечных мутаций были отцовского происхождения.
Различные типы мутаций могут быть переданы от родителя к ребенку или приобретены спонтанно. Механизм, управляющий последним, привлек внимание в последние годы из-за важности этого типа мутации при таких заболеваниях, как шизофрения и аутизм. Скорость мутаций de novo, по-видимому, доминирует с возрастом отца. Скорость здесь возрастает с увеличением возраста отца, возможно, из-за последствий снижения эффективности репликации ДНК или механизмов восстановления, которые, как ожидается, будут ухудшаться с возрастом. Поэтому риск заболевания должен увеличиваться с увеличением возраста отца. Обнаружено, что это происходит во многих случаях, включая синдром Crouzon, множественную эндокринную неоплазию II типа и нейрофиброматоз типа I. Совсем недавно O"Roak et al . наблюдали заметную отцовскую составляющую 51 мутаций de novo, идентифицированных в результате исследования секвенирования 188 родителей-детей со случаями спорадического аутизма. Эти результаты аналогичны тем, которые наблюдаются в последних отчетах о CNN n novo при интелектуальной недостаточности. Эта корреляция может быть объяснена значительно большим числом митотических клеточных делений в зародышевых клетках или сперматоцитах до мейоза в течение всего времени жизни мужчин по сравнению с тем, что происходит при оогенезе у женщин.
Основываясь на установленном числе делений клеток, происходящих в оогенезе (от рождения до менопаузы) по сравнению со сперматогенезом (от полового созревания до конца жизни), Джеймс Ф. Кроу (James F. Crow) подсчитал, что в возрасте 30 лет среднее число повторений хромосомы от зиготы до образование сперматозоидов в 16,5 раз выше, чем от зиготы до образования яйцеклеток.
Генетический мозаицизм обусловлен возникновением de novo митотических мутаций, проявляется очень рано в развитии эмбриона и определяется как наличие множественных клеточных клонов с определенным генотипом у одного и того же человека. Мозаицизм соматической и зародышевой линии существует, но мозаицизм зародышевой линии может способствовать передаче того, что может передаваться мутацией de novo потомству.
Спонтанные мутации, возникающие в соматических клетках (во время митоза, после оплодотворения), также могут играть роль в генезе заболеваний, связанных с нарушениями развития.
Е.В. Тозлиян, педиатр-эндокринолог, генетик, к. м. н., Обособленное структурное подразделение «Научно-исследовательский клинический институт педиатрии» ГБОУ ВПО РНИМУ им. Н.И. Пирогова Минздрава РФ, г. Москва Ключевые слова
: дети, синдром Нунан, диагностика.
Key words
: children, syndrome Noonan, diagnostics.
В статье описан синдром Нунан (синдром Ульриха – Нунан, тернероидный синдром с нормальным кариотипом) – редкая врожденная патология, наследуется по аутосомно-доминантному типу, носит семейный характер, однако встречаются и спорадические случаи. Синдром предполагает наличие фенотипа, характерного для синдрома Шерешевского – Тернера у особей женского и мужского пола с нормальным кариотипом. Представлено клиническое наблюдение. Показаны сложности дифференциальнодиагностического поиска, недостаточная информированность клиницистов о данном синдроме и важность междисциплинарного подхода.
Исторические факты
Впервые о необычном синдроме упомянул О. Kobylinski в 1883 году (фото 1).
Старейший известный клинический случай синдрома Нунан, описан в 1883 году О. Kobylinski
Болезнь описана в 1963 году американским врачом-кардиологом Жаклин Нунан, сообщившей о девяти пациентах со стенозом клапана легочной артерии, малым ростом, гипертелоризмом, умеренным снижением интеллекта, птозом, крипторхизмом и скелетными нарушениями. Доктор Нунан, практиковавшая как детский кардиолог в университете Айовы, заметила, что у детей с редким типом порока сердца – стенозом клапана легочной артерии – часто наблюдались типичные физические аномалии в виде низкого роста, крыловидной шеи, широко посаженных глаз и низко расположенных ушей. Мальчики и девочки поражались одинаково. Доктор Джон Опиц, бывший студент Нунан, первым ввел в употребление термин «синдромом Нунан» для характеристики состояния детей, у которых отмечались признаки, похожие на описанные Нунан. Позже Нунан написала статью «Гипертелоризм с фенотипом Тернера», и в 1971 году на симпозиуме сердечнососудистых заболеваний название «синдром Нунан» стало официально признанным .
Этиология и патогенез
Синдром Нунан представляет собой аутосомно-доминантное заболевание с варьирующей экспрессивностью (рис. 1). Ген синдрома Нунан локализован на длинном плече хромосомы 12 . Не исключена генетическая гетерогенность синдрома. Описаны спорадические и семейные формы синдрома с аутосомно-доминантной формой наследования. В семейных случаях мутантный ген наследуется, как правило, от матери, так как из-за тяжелых пороков развития мочеполовой системы мужчины с этим заболеванием часто бесплодны. Большинство описанных случаев являются спорадическими, вызванными мутациями de novo.
. Аутосомно-доминантный тип наследования
Описанные сочетания синдрома Нунан с нейрофиброматозом I типа в нескольких семьях заставило предположить возможную связь двух независимых локусов 17q11.2 хромосомы 17. У некоторых больных выявляются микроделеции в локусе 22q11 хромосомы 22; в этих случаях клинические проявления синдрома Нунан сочетаются с гипофункцией тимуса и синдромом Ди Джорджи. Ряд авторов обсуждают участие в патогенезе синдрома предполагаемых генов лимфогенеза в связи с наличием сходных с синдромом Тернера лицевых и соматических аномалий и высокой частоты патологии лимфатической системы .
Наиболее частая причина синдрома Нунан – мутация гена PTPN11, которая обнаруживается приблизительно у 50% больных. Белок, кодируемый геном PTPN11, относится к семейству молекул, регулирующих ответ эукариотических клеток на внешние сигналы. Наибольшее число мутаций при синдроме Нунан локализовано в экзонах 3,7 и 13 гена PTPN11, кодирующих домены белка, отвечающие за переход протеина в активное состояние .
Возможные представления о патогенезе представлены следующими механизмами:
RAS-MAPK-путь – очень важный путь сигнальной трансдукции, через который внеклеточные лиганды – определенные факторы роста, цитокины и гормоны – стимулируют клеточную пролиферацию, дифференцирование, выживаемость и метаболизм (рис. 2). После связывания лиганда рецепторы на поверхности клеток фосфорилируются в местах их эндоплазматического региона. Это связывание задействует адаптерные протеины (например, GRB2), которые формируют конститутивный комплекс с факторами обмена гуаниновых нуклеотидов (например, SOS), конвертирующих неактивный ГДФ-связанный RAS в его активную ГТФ-связанную форму. Активированные RAS-протеины затем активируют RAF-MEKERKкаскад через ряд реакций фосфорилирования. В результате активированный ERK проникает в ядро для изменения транскрипции целевых генов и корректирует активность эндоплазматических мишеней для индукции адекватных кратковременных и длительных клеточных ответов на стимул. Все гены, вовлеченные в синдром Нунан, кодируют интегральные для этого пути протеины, и мутации, вызывающие болезнь, обычно усиливают сигнал, проходящий через этот путь.

. RAS-MAPK-сигнальный путь. Ростовые сигналы передаются с активированных фактором роста рецепторов к ядру. Мутации в PTPN11, KRAS, SOS1, NRAS и RAF1 ассоциированы с синдромом Нунан, а мутации в SHOC2 и CBL ассоциированы с подобным синдрому Нунан фенотипом
Клиническая характеристика синдрома Нунан
Фенотип больных с синдромом Нунан напоминает синдром Тернера: короткая шея с крыловидной складкой или низким ростом волос, низкий рост, гипертелоризм глазных щелей (фото 2). Лицевые микроаномалии включают антимонголоидный разрез глазных щелей, опущенные вниз наружные углы глазных щелей, птоз, эпикантус, низко расположенные ушные раковины, складчатый завиток ушных раковин, аномалии прикуса, расщелину язычка мягкого неба, готическое небо, микрогнатию и микрогению. Грудная клетка щитовидной формы с гипоплазированными широко расставленными сосками, грудина выступает в верхней части и западает в нижней. Около 20% больных имеют умеренно выраженную патологию скелета. Наиболее часто встречаются воронкообразная деформация грудной клетки, кифоз, сколиоз; реже – уменьшение числа шейных позвонков и их сращение, напоминающее аномалии при синдроме Клиппеля – Фейля .

. Фенотипы синдрома Нунан
У больных с синдромом Нунан обычно светлые густые вьющиеся волосы с необычным ростом на темени, часто встречаются пигментные пятна на коже, гипертрихоз, дистрофия ногтевых пластинок, аномалии прорезывания и расположения зубов, склонность к образованию келоидных рубцов, повышенная растяжимость кожи. У трети больных отмечаются периферические лимфатические отеки, чаще лимфедема кистей и стоп проявляется у детей раннего возраста. Нередким признаком является патология зрения (миопия, косоглазие, умеренный экзофтальм и др.). Задержка роста встречается примерно у 75% больных, больше выражена у мальчиков и обычно незначительна. Отставание в росте манифестирует в первые годы жизни, реже отмечается незначительный дефицит роста и массы при рождении. С первых месяцев жизни отмечается снижение аппетита. Костный возраст обычно отстает от паспортного.
Характерным признаком синдрома является одно- или двусторонний крипторхизм, встречающийся у 70–75% больных мужского пола, у взрослых больных отмечается азооспермия, олигоспермия, дегенеративные изменения яичек. Тем не менее пубертат наступает спонтанно, иногда с некоторой задержкой. У девочек часто отмечается задержка становления менструации, иногда – нарушения менструального цикла. Фертильность может быть нормальной у больных обоих полов.
Умственная отсталость выявляется более чем у половины больных, как правило, незначительная. Часто отмечаются особенности поведения, расторможенность, синдром дефицита внимания. Речь обычно развита лучше, чем другие интеллектуальные сферы. Степень снижения интеллекта не коррелирует с тяжестью соматических нарушений [Маринчева Г.С., 1988]. В единичных случаях описываются пороки развития центральной нервной системы (гидроцефалия, спинномозговые грыжи), тромбоэмболические инфаркты мозга, возможно, связанные с гипоплазией сосудов .
Пороки внутренних органов при синдроме Нунан достаточно характерны. Наиболее типичными являются сердечно-сосудистые аномалии: клапанный стеноз легочной артерии (около 60% больных), гипертрофическая кардиомиопатия (20–30%), структурные аномалии митрального клапана, дефекты предсердной перегородки, тетрада Фалло; коарктация аорты описана только у больных мужского пола.
У трети больных регистрируются пороки мочевыделительной системы (гипоплазия почек, удвоение лоханок, гидронефроз, мегауретер и др.).
Достаточно часто при синдроме Нунан отмечается повышенная кровоточивость, особенно при оперативных вмешательствах в ротовой полости и носоглотке. Обнаруживаются различные дефекты коагуляции: недостаточность тромбоцитарной системы, снижение уровня факторов свертывания, особенно XI и XII, увеличение тромбопластинового времени . Имеются сообщения о сочетании синдрома Нунан с лейкемией и рабдомиосаркомой, что может свидетельствовать о некотором повышении риска малигнизации у этих больных .
В таблице 1 представлены особенности фенотипа при синдроме Нунан, меняющиеся с возрастом пациента. В таблице 2 – корреляция между фенотипом и генотипом при синдроме Нунан.
Таблица 1 . Типичные черты лица у больных синдромом Нунан по возрастам
| Лоб, лицо, волосы | Глаза | Уши | Нос | Рот | Шея | |
| Новорожденный* | Высокий лоб, низкая линия роста волос в затылочной области | Гипертелоризм, наклонные книзу глазные щели, складка эпикантуса | – | Короткий и широкий утопленный корень, вздернутый кончик | Глубоко утопленный губной желобок, высокие широкие пики красной каймы губ, микрогнатия | Избыточная кожа на затылке |
| Грудной (2–12 мес.) | Большая голова, высокий и выпирающий лоб | Гипертелоризм, птоз или толстые нависающие веки | – | Короткий и широкий утопленный корень | – | – |
| Ребенок (1–12 лет) | Грубые черты, вытянутое лицо | – | – | – | – | – |
| Подросток (12–18 лет) | Миопатичес-кое лицо | – | – | Мостик высокий и тонкий | – | Очевидное формирование шейных складок |
| Взрослый (>18 лет) | Отличительные черты лица утонченные, кожа кажется тонкой и прозрачной | – | – | Выпирающая носогубная складка | – | – |
| Все возрасты | – | Голубые и зеленые радужные оболочки, ромбовидные брови | Низкие, ротированные назад уши с толстыми складками | – | – | – |
Таблица 2 . Корреляции между генотипом и фенотипом при синдроме Нунан*
| Сердечнососудистая система | Рост | Развитие | Кожа и волосы | Другое | |
| PTPN11 (примерно 50%) | Более выражен стеноз легочного ствола; меньше – гипертрофическая кардиомиопатия и дефект межпредсердной перегородки | Более низкий рост; ниже концентрация IGF1 | Пациенты с N308D и N308S имеют слабое снижение или нормальный интеллект | – | Больше выражен геморрагический диатез и ювенильная миеломоноцитарная лейкемия |
| SOS1 (примерно 10%) | Меньше дефект межпредсердной перегородки | Более высокий рост | Меньше снижение интеллекта, задержка развития речи | Подобны сердечно-кожно-лицевому синдрому | – |
| RAF1 (примерно 10%) | Больше тяжелая гипертрофическая кардио-миопатия | – | – | Больше родимых пятен, лентиго, пятен кофе с молоком | – |
| KRAS (<2%) | – | – | Более тяжелая задержка когнитивного развития | Подобны сер-дечно-кожно-лицевому синдрому | – |
| NRAS (<1%) | – | – | – | – | – |
Данные лабораторных и функциональных исследований
Специфических биохимических маркеров для диагностики синдрома Нунан не существует. У некоторых больных выявляется снижение спонтанной ночной секреции гормона роста при нормальном ответе на фармакологические стимулирующие тесты (клофелином и аргинином), снижение уровня соматомедина-С и снижение реакции соматомединов на введение гормона роста.
Критерии диагноза
Диагноз «синдром Нунан» ставится на основании клинических признаков, в некоторых случаях диагноз подтверждается результатами молекулярно-генетического исследования. Критерии диагностики синдрома включают наличие характерного лица (при нормальном кариотипе) в сочетании с одним из следующих признаков: патологии сердца, низкий рост или крипторхизм (у мальчиков), задержка полового созревания (у девочек). Для выявления сердечно-сосудистой патологии необходимо проведение ультразвукового исследования сердца с динамическим определением размеров полостей и стенки желудочков. Возможна пренатальная диагностика заболевания при помощи ультразвукового мониторинга, позволяющего выявить пороки сердца и аномалии строения шеи .
Дифференциальная диагностика
У девочек дифференциальный диагноз проводится в первую очередь с синдромом Тернера; уточнить диагноз позволяет цитогенетическое исследование. Фенотипические признаки синдрома Нунан встречаются при ряде других заболеваний: синдроме Вильямса, синдроме LEOPARD, Дубовица, кардиофацио-кожном синдроме, Корнелии де Ланге, Коэна, Рубинштейна – Тейби и др. Точная идентификация этих заболеваний будет возможна только при проведении молекулярногенетических исследований каждого синдрома при значительном клиническом материале, что в настоящее время активно развивается.
Лечение
Лечение больных с синдромом Нунан направлено на устранение пороков сердечно-сосудистой системы, нормализацию психических функций, стимуляцию роста и полового развития. Для лечения больных с дисплазией клапанов легочной артерии, помимо прочих методов, с успехом применяется баллонная вальвулопластика. С целью стимуляции психического развития применяются ноотропные и сосудистые средства. Препараты, направленные на стимуляцию полового развития, показаны в основном больным с крипторхизмом. Применяются препараты хорионического гонадотропина в возрастных дозировках. В старшем возрасте – при наличии гипогонадизма – препараты тестостерона. В последние годы применяются рекомбинантные формы гормона роста человека в лечении больных с синдромом Нунан . Клинические данные подтверждаются увеличением на фоне терапии уровня соматомедина-С и специфического связывающего белка. Конечный рост больных, длительное время получающих терапию гормоном роста, в некоторых случаях превышает средний рост членов семьи.
Прогноз для жизни определяется тяжестью сердечно-сосудистой патологии.
Профилактика болезни основывается на данных медико-генетического консультирования.
Медико-генетическое консультирование
При медико-генетическом консультировании следует исходить из аутосомно-доминантного типа наследования и высокого (50%) риска повторения заболевания в семье при унаследованных формах. С целью идентификации характера типа наследования необходимо проводить тщательное обследование родителей, так как синдром может проявляться минимальными клиническими симптомами. В настоящее время разработана и совершенствуется молекулярно-генетическая диагностика заболевания путем типирования мутаций в генах: PTPN11, SOS1, RAF1, KRAS, NRAS и др. Разрабатываются способы пренатальной диагностики заболевания.
Клиническое наблюдение
Мальчик Г., 9 лет (фото 3), наблюдался по месту жительства врачом-генетиком с диагнозом «хромосомная патология?, синдром Вильямса (своеобразный фенотип, уплотнение створок митрального клапана, гиперкальциемия однократно в 3 года)?.

. Особенности фенотипа ребенка с синдромом Нунан (удлиненный лицевой скелет с «пухлыми щечками», короткая шея, крыловидные складки на шее, нос укорочен с открытыми вперед ноздрями, пухлые губы, скошенный подбородок, антимонголоидный разрез глазных щелей, неправильный прикус, макростомия)
Жалобы на сниженную память, утомляемость, сниженные темпы роста.
Анамнез семейный : родители русские по национальности, не состоящие в кровном родстве и не имеющие профессиональных вредностей, здоровые. Рост отца – 192 см, рост матери – 172 см. В родословной случаев психических заболеваний, эпилепсии, задержки в развитии не отмечалось.
Анамнез жизни и заболевания : мальчик от 2-й беременности (1-я беременность – м/а), протекавшей с угрозой прерывания на всем протяжении, сопровождающейся многоводием. Роды первые, в срок, стремительные, масса при рождении – 3400 г, длина – 50 см. Закричал сразу, оценка по шкале Апгар – 7/9 баллов. При рождении неонатологом обращено внимание на необычный фенотип ребенка, рекомендовано исследование кариотипа, результат – 46, XY (нормальный мужской кариотип). Был заподозрен врожденный гипотиреоз, проведено исследование тиреоидного профиля, результат – нормальный тиреоидный статус. Далее ребенок наблюдался генетиком с предполагаемым диагнозом «синдром Вильямса». Ранний постнатальный период – без особенностей. Моторное развитие по возрасту, первые слова – к году, фразовая речь – в 2 года 3 мес.
В возрасте 8 лет консультирован эндокринологом по поводу сниженных темпов роста, утомляемости, сниженной памяти. При рентгенологическом исследовании кистей рук выявлено умеренное отставание костного возраста (КВ) от паспортного (КВ соответствовал 6 годам). При исследовании тиреоидного профиля выявлено умеренное повышение тиреотропного гормона при нормальном уровне свободного Т4 и остальных показателей; УЗИ щитовидной железы – без патологии. Назначена гормональная терапия с последующим динамическим наблюдением.
Учитывая неопределенность диагноза по месту жительства, генетиком ребенок направлен в Московский областной консультативно-диагностический центр для детей с целью уточнения диагноза.
Данные объективного исследования:
Рост – 126 см, вес – 21 кг.
Физическое развитие ниже среднего, гармоничное. Sds роста соответствует –1 (норма – –2+2). Особенности фенотипа (фото 3): удлиненный лицевой скелет с «пухлыми щечками», короткая шея, крыловидные складки на шее, низкий рост волос на шее, нос укороченный с открытыми вперед ноздрями, пухлые губы, скошенный подбородок, антимонголоидный разрез глазных щелей, неправильный прикус, макростомия, гипертелоризм сосков, асимметрия грудной клетки, на стопах неполная кожная синдактилия 2–3-го пальцев, выраженная гипермобильность межфаланговых суставов, ломкие, сухие ногти. По внутренним органам – без особенностей. Половое развитие – Tanner I (что соответствует допубертатному периоду).
Данные лабораторных и функциональных исследований:
Клинический анализ крови и мочи – норма.
Биохимический анализ крови – показатели в пределах нормы.
Тиреоидный профиль (ТТГ) – 7,5 мкМЕ/ мл (норма – 0,4–4,0), остальные показатели в норме.
Соматотропный гормон (СТГ) – 7 нг/мл (норма – 7–10), соматомедин-С – 250 нг/мл (норма – 88–360).
УЗИ щитовидной железы – без патологии.
УЗИ внутренних органов – без особенностей.
ЭКГ – синусовая тахикардия, нормальное положение электрической оси сердца.
ЭхоКГ – ПМК I степени с минимальной регургитацией, миксоматозное утолщение створок митрального клапана, дополнительная хорда в полости левого желудочка.
R-графия позвоночника – правосторонний сколиоз грудного отдела позвоночника I степени.
R-графия кистей рук с захватом предплечий – костный возраст 7–8 лет.
ЭЭГ-паттернов эпилептической активности не зарегистрировано.
МРТ головного мозга – без патологических изменений.
Аудиограмма – без патологии.
ДНК-диагностика: молекулярно-генетическое исследование – делеций исследуемых локусов критического района хромосомы 7 не выявлено; обнаружена мутация Gly434Ary (1230G>A) в 11-м экзоне гена SOS1 (анализ гена PTPN11 – мутаций не обнаружено), что характерно для синдрома Нунан.
Консультации специалистов:
Эндокринолог – субклинический гипотиреоз, неполная медикаментозная компенсация.
Окулист – астигматизм.
Невролог – вегетососудистая дистония. Невротические реакции.
Кардиолог – функциональная кардиопатия.
Хирург-ортопед – нарушение осанки. Деформация грудной клетки.
Генетик – синдром Нунан.
Учитывая фенотип ребенка, данные анамнеза, результаты дополнительных исследований, поставлен диагноз «синдром Нунан», что подтверждено результатом молекулярно-генетического исследования.
Таким образом, представленное клиническое наблюдение демонстрирует сложности дифференциально-диагностического поиска, необходимость интегрировать отдельные признаки в общий фенотип того или иного патологического состояния для целенаправленной своевременной диагностики отдельных форм наследственных заболеваний, важность молекулярно-генетических методов для уточнения диагноза. Своевременная диагностика, уточнение генеза каждого синдрома особенно важны, так как позволяют найти оптимальный подход к лечению этих состояний, профилактике возможных осложнений (вплоть до инвалидности ребенка); предупреждению повторного возникновения наследственных болезней в пораженных семьях (медико-генетическое консультирование). Это диктует необходимость врачам различных специальностей четко ориентироваться в потоке наследственно обусловленной патологии.
Список литературы:
- Baird P., De Jong B. Noonan’s syndrome (XX and XY Turner phenotype) in three generations of a family // J. Pediatr., 1972, vol. 80, p. 110–114.
- Hasegawa T., Ogata T. et al. Coarctation of the aorta and renal hupoplasia in a boy with Turner/Noonan surface anomalies and a 46, XY karyotype: a clinical model for the possible impairment of a putative lymphogenic gene(s) for Turner somatic stigmata // Hum. Genet., 1996, vol. 97, р. 564–567.
- Федотова Т.В., Кадникова В.А. и соавт. Клинико-молекулярно-генетический анализ синдрома Нунан. Материалы VI съезда Российского общества медицинских генетиков. Медицинская генетика, приложение к № 5, 2010, с.184.
- Ward K.A., Moss C., McKeown C. The cardio-facio-cutaneous syndrome: a manifestation of the Noonan syndrome? // Br. J. Dermatol., 1994, vol. 131, р. 270–274.
- Municchi G., Pasquino A.M. et al. Growth hormone treatment in Noonan syndrome: report of four cases who reached fi nal height // Horm. Res., 1995, vol. 44, р. 164–167.
Амниоцентез - исследование, которое используется для того, чтобы получить образец для анализа генов и хромосом плода. Плод находится в матке окруженный жидкостью. Эта жидкость содержит небольшое количество клеток кожи будущего ребенка. Небольшое количество жидкости отбирается тонкой иглой через брюшную стенку матери (живот). Жидкость отправляют в лабораторию для исследования. Для получения более подробной информации см. брошюру Амниоцентез.
Аутосомно-доминантное генетическое заболевание - это такое заболевание, для развития которого человеку необходимо унаследовать одну измененную копию гена (мутацию) от одного из родителей. При данном типе наследования заболевание передается половине детей супружеской пары от одного из родителей, который болен. Поражаются оба пола с равной вероятностью. В семьях наблюдается вертикальная передача заболевания: от одного из родителей половине детей.
Аутосомно-рецессивное генетическое заболевание - это такое заболевание, для развития которого человеку необходимо унаследовать две измененные копии гена (мутации), по одной от каждого из родителей. При данном типе наследования болеет четверть детей супружеской пары. Родители здоровы, но являются носителями заболевания. Человек, имеющий только одну копию измененного гена, будет являться здоровым носителем. Для получения более подробной информации см. брошюру Рецессивное наследование.
Аутосомный - признак, ген которого, расположен на аутосомах.
Аутосомы - У человека 23 пары хромосом. Пары от 1 до 22 называются аутосомами и выглядят одинаково у мужчин и женщин. Хромосомы 23-й пары у мужчин и женщин отличаются, и называются половыми хромосомами.
Биопсия ворсин хориона, БВП - процедура, проводимая во время беременности, для забора клеток плода для проведения исследования генов или хромосом будущего ребенка на определенные наследственные состояния. Небольшое количество клеток берется из развивающейся плаценты и отправляется в лабораторию для исследования. Для получения более подробной информации см. брошюру Биопсия ворсин хориона.
Влагалище - орган, соединяющий матку с внешней средой, родовой канал.
Ген - информация, необходимая организму для жизнедеятельности, хранящаяся в химической форме (ДНК) на хромосомах.
Генетический - вызванный генами, имеющий отношение к генам.
Генетическое исследование - исследование, которое может помочь установить, имеются ли изменения в отдельных генах или хромосомах. Для получения более подробной информации см. брошюру Что такое генетическое исследование?
Генетическое заболевание - заболевание, вызванное нарушениями в генах или хромосомах.
Делеция - потеря части генетического материала (ДНК); этот термин можно использовать для обозначения потери части как гена, так и хромосомы. Для получения более подробной информации см. брошюру Хромосомные нарушения.
ДНК - химическая субстанция, из которой состоят гены, и которая содержит информацию, необходимую организму для жизнедеятельности.
Дупликация - аномальное повторение последовательности генетического материала (ДНК) в гене или хромосоме. Для получения более подробной информации см. брошюру Хромосомные нарушения.
Измерение толщины воротникового пространства (ТВП) - ультразвуковое исследование задней части области шеи плода, которое заполнено жидкостью на ранней стадии беременности. Если у ребенка есть врожденное заболевание (например, синдром Дауна), толщина воротникового пространства может быть изменена.
Инверсия - изменение последовательности генов в отдельной хромосоме. Для получения более подробной информации см. брошюру Хромосомные нарушения.
Инсерция - вставка дополнительного генетического материала (ДНК) в ген или хромосому. Для получения более подробной информации см. брошюру Хромосомные нарушения.
Кариотип - описание структуры хромосом индивидуума, включающее число хромосом, набор половых хромосом (XX или XY) и любые отклонения от нормального набора.
Клетка - человеческое тело состоит из миллионов клеток, которые служат «строительными блоками». Клетки в разных местах тела человека по-разному выглядят и выполняют различные функции. Каждая клетка (за исключением яйцеклеток у женщин и сперматозиодов у мужчин) содержит по две копии каждого гена.
Кольцевая хромосома - термин, используемый, когда концы хромосомы соединяются друг с другом и образуют кольцо. Для получения более подробной информации см. брошюруХромосомные транслокации.
Матка - часть тела женщины, в которой во время беременности растет плод.
Медико-генетическое консультирование - информационная и медицинская помощь людям, обеспокоенным наличием в семье состояния, возможно, имеющего наследственную природу.
Мутация - изменение последовательности ДНК определенного гена. Данное изменение последовательности гена приводит к тому, что нарушается содержащаяся в нём информация, и он не может работать правильно. Это может привести к развитию генетического заболевания.
Невынашивание - п реждевременное прекращение беременности, наступившее до того момента, когда ребенок способен выжить вне матки.
Несбалансированная транслокация - транслокация, при которой хромосомная перестройка приводит к приобретению или утрате некоторого количества хромосомного материала (ДНК), либо одновременно к приобретению дополнительного и утрате части исходного материала. Может возникать у ребенка, родитель которого является носителем сбалансированной транслокации. Для получения более подробной информации см. брошюру Хромосомные транслокации.
Носитель хромосомной перестройки - человек, у которого есть сбалансированная транслокация, при которой количество хромосомного материала не уменьшено, и не увеличено, что обычно не вызывает нарушений здоровья.
Носитель - человек, который обычно не страдает заболеванием (в настоящее время), но является носителем одной измененной копии гена. В случае рецессивного заболевания, носитель обычно здоров; в случае доминантного заболевания с поздним дебютом, человек заболеет позднее.
Оплодотворение - слияние яйцеклетки и сперматозоида для создания первой клетки ребенка.
Плацента - орган, прилежащий к внутренней стенкe матки беременной женщины. Плод получает через плаценту питательные вещества. Плацента растет из оплодотворенной яйцеклетки, поэтому она содержит теже гены, что и плод.
Позитивный результат - результат теста, который показывает, что у обследованного человека выявлено изменение (мутация) в гене.
Половые хромосомы - Х-хромосома и Y-хромосома. Набор половых хромосом определяет, является ли индивид мужчиной или женщиной. У женщин - две Х-хромосомы, у мужчин - одна Х-хромосома и одна Y-хромосома.
Предиктивное тестирование - генетическое исследование, направленное на выявление состояния, которое может развиться или разовьется в течение жизни. Когда генетическое исследование направлено на выявление состояния, которое почти неизбежно разовьется в будущем, такое исследование называют пресимптоматическим .
Пренатальная диагностика - исследование, проводимое во время беременности, на наличие или отсутствие генетического заболевания у ребенка.
Реципрокная транслокация - транслокация которая возникает, когда два фрагмента отрываются от двух разных хромосом и меняются местами. Для получения более подробной информации см. брошюру Хромосомные транслокации.
Робертсоновская транслокация - возникает, когда одна хромосома оказывается прикрепленной к другой. Для получения более подробной информации см. брошюру Хромосомные транслокации.
Сбалансированная транслокация - т ранслокация (хромосомная перестройка), при которой количество хромосомного материала не уменьшено и не увеличено, но он перемещен с одной хромосомы на другую. Человек со сбалансированной транслокацией обычно не страдает от этого, однако риск развития генетических заболеваний для его детей повышен. Для получения более подробной иниформации см. брошюру Хромосомные транслокации.
Состояние, сцепленное с полом - См. Х-сцепленное наследование.
Сперматозоид - половая клетка отца, отцовский вклад в образование клетки, из которой разовьется новый ребенок. Каждый сперматозоид содержит 23 хромосомы, по одной из каждой пары хромосом отца. Сперматозоид сливается с яйцеклеткой для создания первой клетки, из которой развивается будущий ребенок.
Транслокация - перестройка хромосомного материала. Возникает, когда фрагмент одной хромосомы отрывается и прикрепляется в другое место. Для получения более подробной информации см. брошюру Хромосомные транслокации.
Ультразвуковое исследование (УЗИ) - безболезненное исследование, при котором звуковые волны используются для создания изображения плода, растущего в матке матери. Оно может проводиться путем перемещения головки сканера по поверхности брюшной стенки (живота) матери или внутри влагалища.
Хромосомы - нитевидные структуры, различимые под микроскопом, которые содержат гены. В норме у человека 46 хромосом. Один комплект из 23 хромосом мы наследуем от матери, второй комплект из 23 хромосом - от отца.
Х-сцепленное заболевание - генетическое заболевание, возникающее в результате мутации (изменения) в гене, расположенном на Х-хромосоме. Х-сцепленные заболевания включают гемофилию, мышечную дистрофию Дюшенна, сидром ломкой Х-хромосомы и множество других. Для получения более подробной информации см. брошюру Х-сцепленное наследование.
ХХ - так обычно представляют набор половых хромосом женщины. В норме у женщины две Х-хромосомы. Каждая из Х-хромосом наследуется от одного из родителей.
Х-хромосома - Одна из половых хромосом. У женщин в норме две Х-хромосомы. У мужчин в норме одна Х-хромосома и одна Y-хромосома.
Яичник/яичники - органы в теле женщины, продуцирующие яйцеклетки.
Яйцеклетка - половая клетка матери, которая послужит основой для создания первой клетки будущего ребенка. Яйцеклетка содержит 23 хромосомы; по одной из каждой пары, имеющейся у матери. Яйцеклетка сливается со сперматозоидом для формирования первой клетки ребенка.
De novo - с очетание из латинского языка, означающее «заново». Используется для описания изменения генов или хромосом (мутаций), которые являются вновь образовавшимися, т.е. ни у одного из родителей человека с мутацией de novo этих изменений нет.
XY - так обычно представляют набор половых хромосом мужчины. В норие у мужчин одна Х-хромосома и одна Y-хромосома. Мужчины наследуют Х-хромосому от матери, а Y-хромосому - от отца.
Y-хромосома - одна из половых хромосом. В норме у мужчин одна Y-хромосома и одна Х-хромосома. У женщины в норме две Х-хромосомы.